
TRUNG TÂM XÉT NGHIỆM ADN LAB SÀI GÒN
ADN ty thể là gì
105
Thứ Hai, 09/11/2023, 03:04 (GMT+7)
ADN ty thể là gì?
ADN ty thể (mtDNA) là ADN có trong ty thể. Loại vật chất di truyền này là một trong những ADN ngoại nhân ở sinh vật nhân chuẩn.
Một dạng ADN khác tồn tại bên ngoài nhân là ADN lục lạp, ADN này lại xuất hiện trong lục lạp.
Tương tự như ADN trong nhân, ADN ngoài nhân như ADN ty thể là một hợp chất hữu cơ được tạo thành từ các chuỗi nucleotide đơn phân tuyến tính. Lần lượt, mỗi thành phần nucleotide được tạo thành từ nhóm photphat, đường deoxyribose và bazơ nitơ.
ADN tham gia vào việc bảo tồn, sao chép và biểu hiện thông tin di truyền.

Lịch sử khám phá ADN ty thể
ADN ty thể lần đầu tiên được xác định và phân lập bởi Margit Nass và Sylvan Nass vào năm 1963, người đã nghiên cứu một số sợi ty thể mà theo hành vi cố định, ổn định và nhuộm màu của chúng, dường như có liên quan đến ADN (Nass & Nass, 1963).
Bằng kính hiển vi điện tử, họ đã tìm thấy axit nucleic bên trong ty thể của phôi gà. Họ công bố phát hiện của mình trên Tạp chí Sinh học Tế bào vào năm 19631. Năm sau, một nhóm nghiên cứu độc lập đã công bố một phát hiện tương tự; họ đã tìm thấy ADN ngoại nhân trong ty thể của nấm men2. Vì ADN ngoại nhân được tìm thấy trong ty thể nên axit nucleic này cuối cùng được gọi là ADN ty thể để xác định nó với ADNA trong nhân.
Tuy nhiên, trình tự hoàn chỉnh của mtDNA đầu tiên chỉ được công bố và thiết lập dưới dạng Trình tự tham chiếu mtDNA Cambridge (CRS) mười tám năm sau, vào năm 1981 (Anderson và cộng sự, 1981).
Đặc điểm ADN ty thể
ADN ty thể khác biệt về mặt di truyền với ADN trong nhân.
Toàn bộ thông tin di truyền trong ty thể được gọi là hệ gen ty thể hoặc hệ gen nguyên phân.
Ở hầu hết các sinh vật nhân chuẩn đa bào, bộ gen nguyên phân được tổ chức dưới dạng ADN vòng trong khi ở hầu hết các sinh vật nhân chuẩn đơn bào, bộ gen nguyên phân được tổ chức dưới dạng ADN tuyến tính.
Ở động vật có vú, ADN ty thể là ADN vòng kép. Một trong hai sợi rất giàu guanine và được gọi là sợi nặng (hoặc sợi H). Sợi còn lại rất giàu cytosine và được gọi là sợi nhẹ (hoặc sợi L). Chuỗi H được biết là mã hóa 9 gen trong khi chuỗi L mã hóa 28 gen, do đó, có tổng số 37 gen.
Hệ gen nguyên phân của động vật có vú mã hóa cho 13 protein, 22 tRNA và 2 cho rRNA. Vì ty thể có vật liệu di truyền riêng và có khả năng tự sản xuất RNA và protein nên nó được cho là một cấu trúc tế bào chất bán tự trị, tự sinh sản.
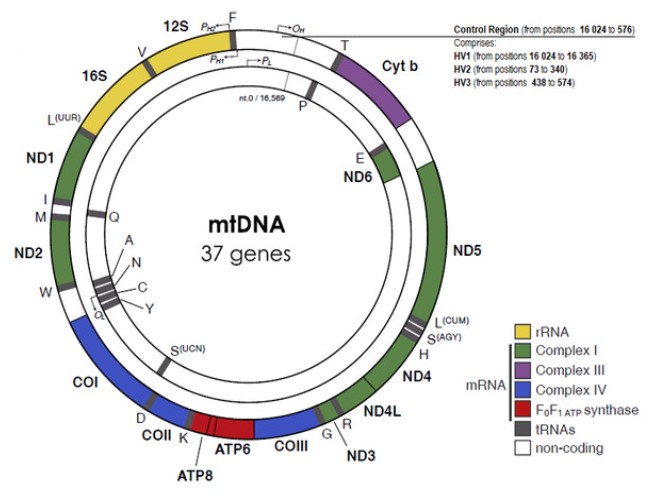
Về cơ bản, mtDNA là một phân tử ADN sợi đôi hình tròn 5 mm không có histone, với khoảng 16 569 cặp bazơ và trọng lượng 107 Dalton (Taanman, 1999). Các chuỗi mtDNA có mật độ khác nhau do thành phần bazơ G+T khác nhau.
Chuỗi nặng (H) mã hóa nhiều thông tin hơn, với các gen cho hai rRNA (12S và 16S), 12 polypeptide và 14 tRNA, trong khi chuỗi nhẹ (L) mã hóa 8 tRNA và một polypeptide.
Tất cả 13 sản phẩm protein đều là một phần của phức hợp enzyme tạo thành hệ thống phosphoryl hóa oxy hóa. Các đặc điểm đặc trưng khác của mtDNA là các gen không có gen và các trình tự giữa các thế hệ bị hạn chế, hoặc thậm chí không có, ngoại trừ ở một vùng quy định.
Vòng D ty thể là vùng ba sợi được tìm thấy trong vùng không mã hóa chính (NCR) của nhiều bộ gen ty thể và được hình thành bằng cách kết hợp ổn định chuỗi DNA 680 bazơ thứ ba được gọi là DNA 7S (Kefi-Ben Atig et cộng sự, 2009).
Điểm gốc của sự sao chép nằm ở vùng không mã hóa hoặc vùng D-loop, đoạn 1 121 cặp bazơ nằm giữa các vị trí 16 024 và 576, theo cách đánh số CRS (Anderson và cộng sự, 1981).
Vùng D-loop cũng bao gồm hai yếu tố thúc đẩy phiên mã, một cho mỗi chuỗi.
Vị trí nucleotide trong bộ gen mtDNA được đánh số theo quy ước được trình bày bởi Anderson et al. (1981), được sửa đổi một chút bởi Andrews et al. (1999), xác định việc thay thế CRS cho rCRS (Trình tự tham chiếu Cambridge đã sửa đổi).
Chính xác hơn, việc ký hiệu bằng số của mỗi cặp bazơ được bắt đầu ở một vị trí tùy ý trên chuỗi H, tiếp tục sau đó và xung quanh phân tử cho khoảng 16 569 cặp bazơ.
Đặc điểm sinh học di truyền của ADN ty thể
Việc thiếu cơ chế sửa chữa mtDNA rõ ràng và độ chính xác thấp của mtDNA polymerase dẫn đến tỷ lệ đột biến cao hơn đáng kể trong bộ gen của ty thể khi so sánh với bộ gen hạt nhân.
Hầu hết các biến thể trình tự giữa các cá thể được tìm thấy ở hai phân đoạn cụ thể của vùng kiểm soát, cụ thể là ở vùng siêu biến 1 (HV1, vị trí 16 024 đến 16 365) và ở vùng siêu biến 2 (HV2, vị trí 73 đến 340) (Greenberg , Newbold & Sugino, 1983). Vùng siêu biến thứ ba (HV3, các vị trí từ 438 đến 574), với các vị trí đa hình bổ sung có thể hữu ích trong việc phân giải các mẫu HV1/HV2 không thể phân biệt được (Lutz và cộng sự, 2000). Kích thước nhỏ và tính biến thiên tương đối cao giữa các cá nhân của vùng HV là những đặc điểm rất hữu ích cho mục đích thử nghiệm pháp y.
Trình tự mtDNA xác định kiểu đơn bội riêng lẻ được báo cáo bởi các cặp bazơ khác nhau liên quan đến trình tự mtDNA rCRS. Tập hợp các kiểu đơn bội tương tự được xác định bằng sự kết hợp của các đa hình nucleotide đơn (SNP) trong mtDNA được thừa hưởng từ một tổ tiên chung xác định một nhóm đơn bội được hình thành do sự tích lũy tuần tự các đột biến thông qua dòng mẹ (Mitchell và cộng sự, 2015).
Một ty thể chứa từ 2 đến 10 bản sao mtDNA và mỗi tế bào soma có thể có tới 1.000 ty thể (Elson và cộng sự, 2001; Wei và cộng sự, 2017). Do đó, khi số lượng DNA được chiết xuất khá nhỏ hoặc bị phân hủy, có nhiều khả năng thu được kết quả phân loại DNA bằng cách gõ mtDNA hơn là gõ các vùng đa hình được tìm thấy trong DNA hạt nhân.
Ngược lại với DNA nhân, mtDNA chỉ được di truyền từ mẹ sang các con (di truyền theo dòng mẹ), điều này chứng minh thực tế rằng, ngoài đột biến, trình tự mtDNA của anh chị em ruột và tất cả họ hàng bên ngoại đều giống hệt nhau (Case & Wallace, 1981; Giles et al., 1980; Hutchison et al. ., 1974).
Đặc điểm cụ thể này có thể rất hữu ích trong các trường hợp pháp y, chẳng hạn như trong phân tích hài cốt của một người mất tích, trong đó họ hàng bên ngoại có thể cung cấp một số mẫu tham chiếu để so sánh trực tiếp với loại mtDNA. Do thiếu sự tái tổ hợp, họ hàng ngoại từ nhiều thế hệ ngoài nguồn bằng chứng (hoặc vật liệu sinh học) có thể được sử dụng làm mẫu tham chiếu (Case & Wallace, 1981; Giles et al., 1980; Hutchison et al., 1974).
Bản chất đơn bội và đơn dòng của mtDNA ở hầu hết các cá nhân giúp đơn giản hóa quá trình giải thích kết quả giải trình tự DNA. Tuy nhiên, đôi khi vẫn có thể tìm thấy dị thể trong một số trường hợp (Bendall, Macaulay & Sykes, 1997; Bendall & Sykes, 1995; Comas, Paabo & Bertranpetit, 1995; Gill và cộng sự, 1994; Ivanov và cộng sự, 1996; Wilson và cộng sự cộng sự, 1997).
Một người được coi là dị thể nếu người đó mang nhiều hơn một loại mtDNA có thể phát hiện được.
Có hai loại dị hợp tử, liên quan đến đa hình chiều dài và thay thế điểm. Chỉ có điều sau là quan trọng để nhận dạng con người pháp y.
Hầu hết các phòng thí nghiệm pháp y trên toàn thế giới không báo cáo tính đa hình về chiều dài và các hướng dẫn về nhận dạng con người bằng mtDNA không coi chúng là thông tin bắt buộc (Parson và cộng sự, 2014; Prinz và cộng sự, 2007). Hơn nữa, thông tin về đa hình độ dài không có tác động gì đến định nghĩa của nhóm đơn bội.
Dị thể biểu hiện theo nhiều cách khác nhau (Stewart và cộng sự, 2001). Một cá nhân có thể biểu hiện nhiều loại mtDNA trong một mô. Một cá thể có thể là dị chất trong một mẫu mô và đồng chất trong một mẫu mô khác. Cuối cùng, một cá nhân có thể biểu hiện một loại mtDNA trong một mô và một loại khác trong một mô khác. Trong ba tình huống có thể xảy ra, tình huống cuối cùng ít có khả năng xảy ra nhất. Khi dị hợp tử được tìm thấy trong mtDNA của một cá thể, nó thường khác nhau ở một bazơ duy nhất, ở HV1 hoặc HV2.
Dị thể được quan sát thấy ở vị trí 16.169 của vùng kiểm soát mtDNA trong hài cốt giả định của Sa hoàng Nicholas II của Nga và anh trai ông, Đại công tước Nga Georgij Romanov (Gill và cộng sự, 1994; Ivanov và cộng sự, 1996). Lần lượt, Comas, Paabo & Bertranpetit (1995) đã phát hiện dị thể ở hai vị trí riêng biệt, 16, 293 và 16.311, trong mtDNA của mái tóc được nhổ của một người hiến tặng ẩn danh. Wilson và cộng sự. (1997) đã tìm thấy một gia đình gồm có một người mẹ và hai đứa con mang mtDNA dị thể ở vị trí 16.355 cả trong mẫu máu và mẫu phết niêm mạc miệng.
Sự tồn tại của các cá thể dị thể và kiến thức hạn chế về cả cơ chế cũng như tỷ lệ dị thể có thể là vấn đề được nêu ra trong nỗ lực loại trừ bằng chứng mtDNA khỏi các cuộc điều tra pháp y. Dị thể ở một vị trí nucleotide được quan sát thường xuyên hơn trong các mẫu tóc, chủ yếu là do sự trôi dạt di truyền và do tắc nghẽn xảy ra do tính chất bán dòng của nang lông (Budowle et al., 200).




